

The Astonishing Preservation of Dinosaur Soft Tissue: Evidence of a Recent Flood Catastrophe
- Dr. Robert L. Wright

- Jun 20
- 11 min read
This investigation began while on a dig with Dr. Mark Armitage in Glendive, Montana, in 2020. As we excavated and discussed the specimens before us, we were constantly amazed at the extraordinary level of preservation we witnessed. These bones did not look thousands of years old, let alone millions or hundreds of millions of years old. What mechanism could possibly explain their existence? How could soft tissues remain intact after all this supposed time? What had God accomplished during the Flood that allowed us to witness this remarkable evidence today? After several years of study, I am confident I have some of the answers. They fill me with awe of the Master Chemist, the Architect of all Creation, our God.
In the world of paleontology, few discoveries have challenged long-held assumptions as profoundly as the recovery of soft tissues from dinosaur bones. Flexible blood vessels, stretchy fibrous matrices, elastic sheets of fibrillar bone, and even what appear to be intact cells, peripheral nerves, and blood remnants have been extracted from specimens long assumed to be tens of millions of years old. These finds are not mere curiosities. They stand as powerful testimony to the truth of Scripture. The bones of these ancient creatures cannot possibly be hundreds of millions or even millions of years old. Instead, they point unmistakably to burial and preservation during the global Flood described in Genesis, just thousands of years ago.
Paleontologist Mary Schweitzer and her team first reported these remarkable finds in 2005, pulling supple, elastic blood vessels and osteocyte-like cells from the femur of a Tyrannosaurus rex. Building on this, researcher and microscopist Dr. Mark Armitage made groundbreaking contributions through his work at the Hell Creek Formation in Montana. In 2012, Dr. Armitage and colleague Kevin Anderson excavated a large Triceratops horridus supraorbital horn. Upon microscopic examination, Armitage discovered thin, elastic, reddish-brown sheets of soft fibrillar bone tissue that could be peeled away and stretched significantly. Decalcification revealed osteocytes, blood vessels, and other delicate structures. Dr. Armitage has continued this research through the Dinosaur Soft Tissue Research Institute, publishing further firsts such as the identification of peripheral nerves in Triceratops occipital condyle bone. These structures match modern vertebrate nerves in microscopic detail and retain flexibility.
Dr. Armitage’s work extends beyond Cretaceous dinosaurs. He has also reported soft tissue preservation in Permian fossils from Oklahoma, dated conventionally to around 290 million years old. These include blood clots preserved within vascular canals of amphibian limb bones (such as those of Eryops and Cacops) and exceptionally clear peripheral nerve segments showing diagnostic connective tissue patterns. Such discoveries in even what is conventionally considered older strata further underscore the inconsistency of deep-time assumptions with the observed biological material.
Subsequent work has revealed similar preservation in other specimens: collagen proteins, potential red blood cell remnants, and transparent, branching vessels that still respond to chemical treatments much like fresh tissue. Conventional evolutionary thinking insists these dinosaurs roamed the earth 65 to 68 million years ago (and Permian creatures far earlier). Yet biology and chemistry tell a different story. Organic materials like proteins, collagen, blood components, and nerves degrade rapidly under normal conditions. Experiments show that even in ideal laboratory storage, soft tissues break down over centuries, not eons. The persistence of these delicate structures demands rapid burial followed by conditions that halted decay almost immediately.

This aligns perfectly with the biblical account of Noah’s Flood. A cataclysmic event involving the bursting of the fountains of the great deep and the windows of heaven. Vast tectonic upheavals, volcanic activity, and massive sediment flows would have entombed countless creatures in a matter of days or weeks. The Flood was not a gentle rain but a violent, earth-shaping judgment that mixed waters, minerals, and sediments on a global scale. In such chaos, organisms from diverse ecosystems, spanning what we now call Permian to Cretaceous, were swept up, rapidly buried, and subjected to unique geochemical environments that promoted preservation rather than dissolution.
A Pervasive Mechanism for Preservation Across Ecosystems
Fossils, while not as overwhelmingly abundant as one might expect from a pre-Flood world teeming with life, are still remarkably common and widespread. This distribution requires a mechanism that operated pervasively across marine, terrestrial, and freshwater environments. Not every creature was perfectly preserved. Many were disarticulated or destroyed by the violence of the waters. Yet enough survived in remarkable condition to form the fossil record we see today. The key lies in the Flood’s mineral-rich, superheated waters.
As the fountains of the great deep erupted, they released enormous volumes of water saturated with dissolved minerals from the earth’s interior. Volcanic and hydrothermal inputs would have further loaded these waters with silica, iron, carbonates, and other compounds. Superheated conditions under pressure could accelerate mineral precipitation while simultaneously inhibiting bacterial decay. Rapid burial in fine sediments excluded oxygen, creating anoxic microenvironments. Together, these factors created a perfect storm for both mineralization and the stabilization of remaining organic components.

Sodium Silicate: A Key Agent in Rapid Preservation
I propose a key agent in the preservation process is sodium silicate (Na₂SiO₃). In alkaline, silica-rich solutions, conditions likely prevalent as Flood waters interacted with volcanic ash, dissolved silicates from crustal materials, and hydrothermal fluids, sodium silicate readily forms and can penetrate tissues rapidly. Laboratory experiments and natural analogs demonstrate its effectiveness in replicating the types of preservation seen in dinosaur and Permian fossils, including delicate structures rich in proteins, carbohydrates, and especially lipids.

The detailed chemical process unfolds as follows. During the intense Flood dynamics, massive dissolution of silica from volcanic ash, rhyolitic materials, and silicate minerals elevated concentrations of silicic acid (H₄SiO₄) and sodium silicate species in the supersaturated, alkaline waters. These solutions infiltrated the highly porous structure of bones, cartilage, and associated soft tissues through capillary action, pressure gradients from rapid burial, and fluid flow. Bone and vascular porosity, combined with the elevated temperatures and pressures in the Flood environment, facilitated deep penetration before significant decomposition could occur.

Once inside the cellular and vascular spaces, the alkaline sodium silicate solution encountered localized pH shifts triggered by the release of organic acids from initial autolysis and microbial activity. These pH drops, often moving from alkaline toward neutral or slightly acidic ranges, destabilized the monomeric and oligomeric silicate species. This triggered rapid polymerization: silicate ions (SiO₄⁴⁻ and related species) condensed via siloxane (Si-O-Si) bond formation, building chains, clusters, and eventually a three-dimensional network. The result is a viscous silica sol (colloidal silica) that transitions into a hydrogel. This gel fills intercellular spaces, coats cell walls, osteocyte lacunae, and blood vessel linings, and impregnates collagenous and fibrillar matrices.

The silica hydrogel provides multiple protective functions critical for the survival of a wide array of soft tissues. It physically encapsulates biomolecules, excluding oxygen, excess water, and microbial enzymes. Hydrogen bonding between silanol groups (Si-OH) and organic functional groups (such as hydroxyls on collagen or polysaccharides) stabilizes proteins against hydrolysis and denaturation. In experimental settings, this process replicates fine histological details, including cell outlines and fibrous structures, consistent with the elastic sheets and nerve tissues recovered by Dr. Armitage. Dehydration and aging of the gel lead to syneresis (shrinkage and water expulsion) followed by recrystallization into more stable phases such as opal-A, opal-CT, or microcrystalline quartz. This produces permineralized yet often flexible composites when the mineral phase is later partially removed in laboratory decalcification.

This mechanism is particularly vital for the preservation of lipids, which face some of the most severe challenges among soft-tissue components. Lipids, including phospholipids in cell membranes, cholesterol, and fatty acids, are highly susceptible to rapid degradation through hydrolysis (breakdown by water into fatty acids and glycerol), oxidation (especially of polyunsaturated fatty acids, leading to rancidity and chain reactions that fragment molecules), and microbial attack by lipases. In normal burial environments, these processes cause lipid membranes to disintegrate within years to decades, leaving little to no trace. The hydrophobic nature of many lipids makes them prone to leaching or emulsification in aqueous settings, while free radicals accelerate peroxidation, destroying structural integrity. Without intervention, the delicate bilayer structure of cell membranes and intracellular lipid droplets should not persist even over thousands of years, let alone the supposed millions.

The silica hydrogel addresses these vulnerabilities directly by creating a sealed, anoxic, and dehydrated microenvironment. The gel coats and impregnates lipid-rich structures, forming a physical and chemical barrier that limits water access (reducing hydrolysis), blocks oxygen (preventing oxidative chain reactions), and excludes microbes and their enzymes. Hydrogen bonding and physical encapsulation further stabilize lipid molecules, preventing fragmentation and leaching. This explains the recovery of intact membrane remnants and other lipid-derived structures alongside proteins and nerves in specimens studied by Schweitzer and Armitage. Modern experiments with silica solutions on organic tissues confirm the ability to preserve fine cellular details, including membrane-like components, with high fidelity.
Modern experiments also validate the speed and fidelity of this mechanism overall. Wood and organic tissues soaked in sodium metasilicate or silicic acid solutions can achieve significant silicification in days to weeks, with full petrifaction-like results in months under accelerated conditions. Hot spring studies show wood silicifying to 40 percent by weight in just seven years in silica-rich waters, while laboratory simulations using ethyl silicate or sodium silicate solutions replicate cellular structures with high precision. In Flood-scale conditions, with vastly higher mineral loads, temperature fluctuations, and sediment dynamics, the process would have operated orders of magnitude faster and more pervasively.
This silica-based sealing explains the permineralization observed in many specimens, including Dr. Armitage’s Triceratops horn tissues and his Permian amphibian bones, while preserving the elastic properties of the remaining organic cores. The resulting silica gel acts as a robust barrier against further decay, consistent with the stretchable yet stabilized nature of the recovered tissues.
Supporting evidence for silica involvement comes from direct analyses of fossil bones. In some Cretaceous bird fossils, preserved chondrocytes (cartilage cells) were infused with aluminum and silica rather than iron, demonstrating that silica compounds can play a key role in stabilizing soft structures. Demineralized dinosaur bone vessels and fibrous remnants often show silicon-dominated compositions under energy-dispersive X-ray spectroscopy (EDS), with quartz crystals and silica-rich infillings present alongside them. Silica concretions form readily in environments where organic decay lowers pH in silica-rich pore waters, a process easily scaled up in the Flood’s turbulent, mineral-laden conditions. These observations lend strong credibility to a silica-based sealing mechanism operating pervasively across ecosystems.
Iron as a Preservative: Significant Limitations
Some researchers, including Mary Schweitzer, have proposed that iron released from hemoglobin could serve as a natural preservative. In this model, iron from decomposing red blood cells participates in Fenton-type chemistry, generating hydroxyl radicals that create cross-links between proteins such as collagen. These cross-links, along with iron oxyhydroxide precipitates, are said to stabilize tissues and inhibit microbial decay. In the Flood context, iron mobilized from massive erosion and volcanic activity could have infused remains quickly, potentially acting as a fixative in localized contexts.
However, this hypothesis faces substantial challenges as a primary or sufficient mechanism for the observed preservation and is not viable for explaining tissues that have remained flexible and biologically authentic over supposed deep time.
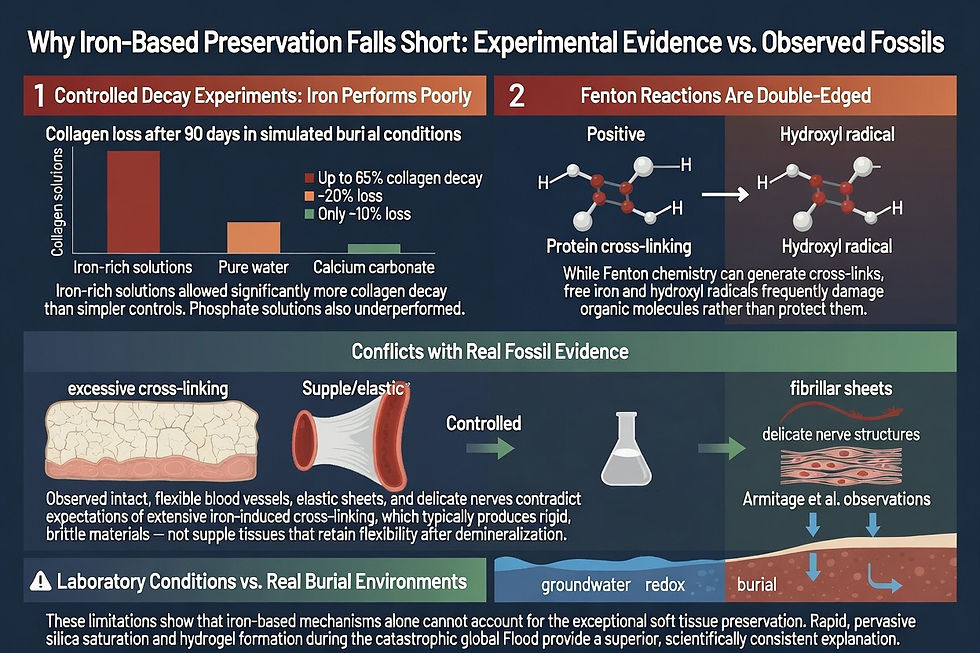
Other potential agents released in the turbulent mixing of ecosystems include carbonates, phosphates, or aluminum compounds, many of which are routinely detected in fossil bone matrices through recrystallization and permineralization. The result: bones that appear millions of years old by flawed radiometric assumptions but contain tissues inconsistent with deep time.
Critics of a young-earth view propose exotic mechanisms to stretch preservation over millions of years. Yet these stretch credibility when weighed against observable decay rates. The simplest and most biblically consistent explanation is rapid, catastrophic burial thousands of years ago. The Flood provides the scale, energy, and chemistry needed. Dinosaur (and Permian) soft tissues, as documented by researchers like Schweitzer and Armitage, are not anomalies. They are expected remnants of a world overwhelmed by judgment and then reshaped by God’s mercy.
Implications for Faith and Science
The pervasive preservation of soft tissues across diverse fossil specimens carries profound implications that reach far beyond laboratory curiosity. It directly upends the assumption of deep time for fossils. Organic decay rates are well-established: proteins such as collagen have half-lives measured in thousands of years at best under ideal conditions, and far shorter in most burial environments. The presence of flexible vessels, intact cells, stretchable fibrillar bone, peripheral nerves, and blood-like remnants in specimens from Permian through Cretaceous layers is chemically and biologically incompatible with tens or hundreds of millions of years. These materials should have long since degraded into indistinguishable mineral replacements or vanished entirely. Their persistence demands rapid entombment and immediate geochemical stabilization, precisely the conditions provided by the global Flood.

This evidence powerfully supports the Flood narrative in Genesis. The catastrophic mixing of ecosystems, superheated mineral-rich waters from the fountains of the great deep, volcanic inputs, and rapid burial in oxygen-poor sediments created the exact geochemical perfect storm needed for silica saturation and other mineral sealing processes. What we observe is not isolated anomalies but a widespread pattern consistent with a single, recent, globe-engulfing event. The fact that soft tissue preservation appears across multiple ages and taxa, from dinosaurs to Permian amphibians, demonstrates the pervasive, uniform action of one massive watery catastrophe rather than millions of years of separate, localized burials.

These discoveries confirm the accuracy and reliability of God’s record in Scripture. The Bible presents a young earth, a historical global Flood as divine judgment, and a subsequent reshaping of the world. Dinosaur and Permian soft tissues stand as physical witnesses crying out from the rocks in agreement with that history. They affirm that the biblical timeline is not metaphorical but literal and trustworthy. Just as Noah’s Ark preserved a remnant through the waters, the Flood itself preserved these biological treasures as testimony for future generations.
For non-biblical paleontologists committed to evolutionary deep time, these findings represent a growing embarrassment and a serious scientific crisis. Initial reports were met with skepticism and accusations of contamination, yet repeated confirmations, from Schweitzer’s T. rex to Armitage’s Triceratops nerves and Permian blood clots, have made dismissal untenable. As more specimens from additional animals and ages reveal the same delicate structures, the challenge intensifies. Researchers must either invoke increasingly ad hoc, untestable mechanisms to preserve organics for eons or confront the collapse of the millions-of-years framework. Many respond with silence, redefinition of terms, or appeals to unknown exceptional conditions, yet none adequately explain the data within a deep-time paradigm. The soft tissue revolution is quietly undermining the foundational assumption of uniformitarian geology and evolutionary biology.
In the end, dinosaur soft tissue is one of the strongest physical evidences for a recent creation and global Flood. It declares the glory of God, exposes the fragility of man-centered science, and calls believers to renewed confidence in Scripture. As we examine the fossil evidence, including the remarkable Triceratops specimens and Permian bones studied by Dr. Armitage, we see not random death but ordered testimony: a young earth, a global catastrophe, and a Creator who remembers mercy in the midst of judgment. Dinosaur soft tissue, sealed by the very minerals stirred up in the Flood, is one more voice crying out from the rocks, declaring the glory of God and the truth of His Word.
References
Armitage, Mark H. "First Report of Peripheral Nerves in Bone from Triceratops horridus Occipital Condyle." Microscopy Today, 2021. Available at Dinosaur Soft Tissue Research Institute (DSTRI): https://dstri.org.
Armitage, Mark H., and Kevin L. Anderson. "Soft Sheets of Fibrillar Bone from a Fossil of the Supraorbital Horn of the Dinosaur Triceratops horridus." Acta Histochemica 115, no. 6 (2013): 603–608.
Armitage, Mark H. Various reports on Permian soft tissue (e.g., blood clots in Eryops and Cacops vascular canals and peripheral nerves), Dinosaur Soft Tissue Research Institute publications and updates, 2020–2023.
Akahane, H., et al. "Rapid Wood Silicification in Hot Spring Water: An Explanation of Silicification of Wood During the Earth's History." Sedimentary Geology 169 (2004): 219–228.
Ballhaus, C., et al. "The Silicification of Trees in Volcanic Ash: An Experimental Study." Geochimica et Cosmochimica Acta 84 (2012): 62–74.
Boatman, Elizabeth M., et al. "Mechanisms of Soft Tissue and Protein Preservation in Tyrannosaurus rex." Scientific Reports 9 (2019).
Chen, G.C. "Treatment of Wood with Polysilicic Acid Derived from Sodium Silicate for Fungal Decay Protection." USDA Forest Service Research (2009).
Drum, R.W. "Silicification of Wood." Botanical studies using sodium metasilicate solutions (1968).
Demassa, T., and Boudreaux, D. "Dinosaur Peptide Preservation and Degradation." Creation Research Society Quarterly (2015).
Götze, J., et al. "Silicification of Wood in the Laboratory." Ceramics – Silikáty 52, no. 4 (2008): 268–277.
Hicks, Hamilton. "Mineralized Sodium Silicate Solutions for Artificial Petrification of Wood." United States Patent US4612050A, September 16, 1986.
Leo, R.F., and E.S. Barghoorn. "Silicification of Wood." Botanical Museum Leaflets (Harvard University) 25 (1976).
Peng, Y., Han, Y., and Gardner, D.J. "Sodium Silicate Coated Wood." Proceedings of the International Convention of Society of Wood Science and Technology and United Nations Economic Commission for Europe, Timber Committee, Geneva, October 11–14, 2010.
Peng, Y., Han, Y., and Gardner, D.J. "Southern Pine Impregnated with Silicate Solution Containing Cellulose Nanofibrils." Holzforschung 66 (2012): 521–527.
Schweitzer, Mary H., et al. "Soft-Tissue Vessels and Cellular Preservation in Tyrannosaurus rex." Science 307, no. 5717 (2005): 1952–1955.
Schweitzer, Mary H., et al. "Soft Tissue and Cellular Preservation in Vertebrate Skeletal Elements from the Cretaceous to the Present." Proceedings of the Royal Society B 274 (2007): 183–197.
Senter, Phil J. "Soft Tissues in Fossil Bone." Palaeontologia Electronica 25, no. 1 (2022).
Ullman, et al. (reported in Sarfati, Jonathan. "Iron Fails to Preserve Collagen." Creation 46, no. 4 (2024)).



Comments